

Por Rosana Souza-Lima
A Terra tem cerca de 4,5 bilhões de anos, e os primeiros fósseis possuem cerca de 3,5 bilhões de anos. Imagina-se que aos poucos foi se acirrando a competição pelas moléculas orgânicas disponíveis para nutrição, e há cerca de 3,4 bilhões de anos surgiram pequenos organismos unicelulares que conseguiam usar substâncias inorgânicas simples para produzir a energia necessária às suas atividades. Esse foi o caminho que permitiu que prosseguisse a vida na Terra – a quebra da molécula de H2O, que era abundante na terra, liberando O2 que passou a ser utilizado pela maioria dos organismos autotróficos.
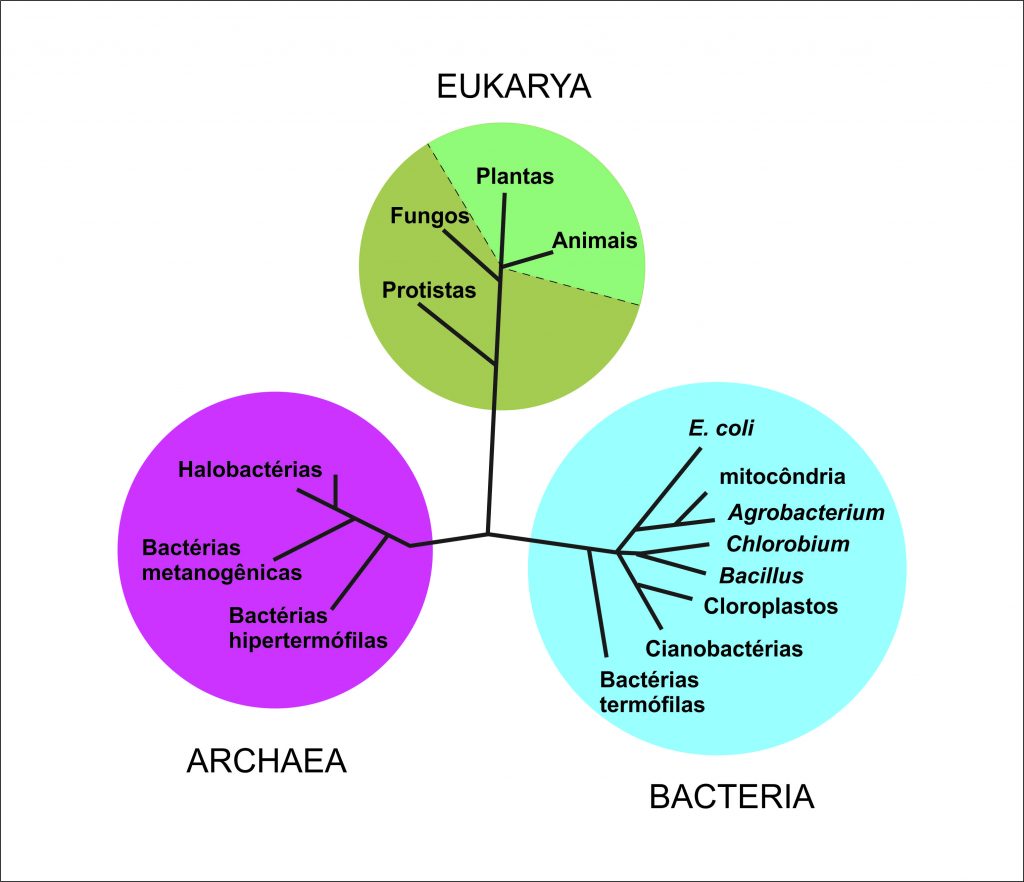
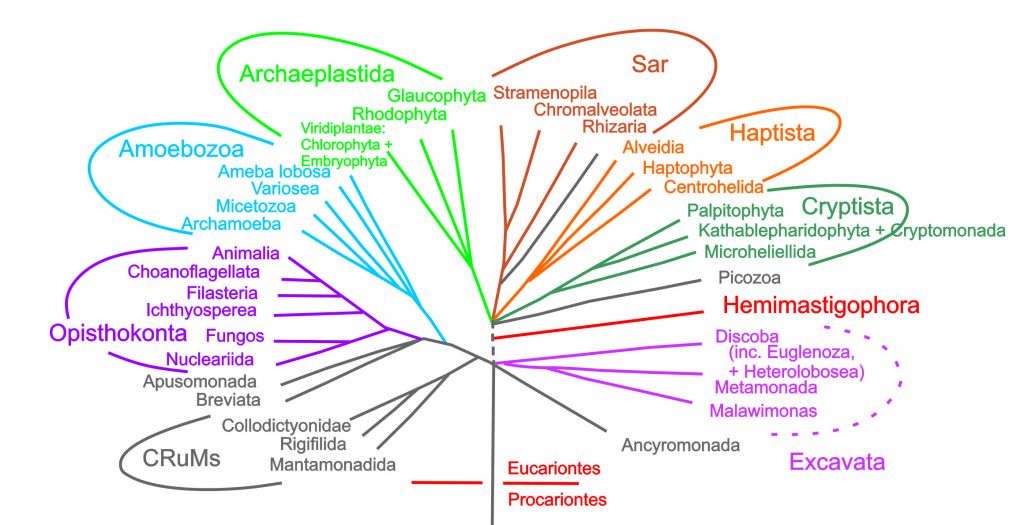
Em 1990, o microbiologista Carl Woese (1928-2012) propôs o agrupamento dos seres vivos em três domínios: dois constituídos por procariontes (Bacteria e Archaea), e um terceiro reunindo todos os seres eucariontes, Eukarya (Figura 1). Uma das mais corroboradas propostas de divisão do Domínio Eukarya é a que une no grupo Opisthokonta os dois mais conhecidos grupos de Eukarya heterótrofos: os fungos e os animais. Opisthokonta é definido pelo fato da maioria das células de seus membros apresentar cristas mitocondriais planas, e as células flageladas normalmente terem um único flagelo originando-se na extremidade posterior da célula (Figura 2).
Há evidências de que a vida procarionte (que são as células com material genético solto no citoplasma) tenha surgido nos oceanos primitivos e há cerca de 2 bilhões de anos, uma dessas células tornou-se endossimbionte (= passou a viver dentro) de outra: isso teria originado as mitocôndrias, resultando numa célula eucariótica que tinha a possibilidade de lidar com a oferta de oxigênio, que estava começando a aparecer na atmosfera da Terra. A célula eucariótica é a que tem o material genético guardado dentro de uma bolsinha membranosa, o núcleo. Milhares de anos se passaram desde a origem desses primeiros organismos eucariontes até eles de fato se diversificarem, mas talvez essa lacuna tenha sido criada por esses organismos terem corpo pequeno e sem esqueletos duros, dificultando que deixassem registro fóssil. Aparentemente ainda não havia oxigênio suficiente para permitir a evolução de formas maiores de microrganismos e, na verdade, nem todo organismo se torna um fóssil, ao morrer: são necessárias condições especiais para que isso aconteça (falamos sobre isso em “O que são fósseis, como se formam e sua importância”).
Atualmente estima-se que os primeiros animais teriam menos de 900 Ma (Ma = milhões de anos). Vários desses registros são icnofósseis, vestígios deixados nas rochas pelas atividades realizadas por esses organismos. Embora haja vestígios até mesmo de formas embrionárias, ainda há muita discussão sobre a idade dos primeiros bilatérios, que parece girar em torno de 565 milhões de anos, na era Neoproterozóica (Pré-Cambriano). Aparentemente o grupo-irmão dos animais são “protistas” coanoflagelados, organismos geralmente coloniais, formados por uma única célula e com um único flagelo, circundado por um colar transparente contendo filamentos de actina. A proposta é de que uma linhagem colonial tenha diferenciado células corporais e reprodutivas, levando ao surgimento de organismos multicelulares cujas células são mantidas juntas por estruturas adesivas: os animais! Outras linhagens de “protistas” foram as ancestrais de Fungos e Plantas, grandes grupos de organismos pluricelulares.
Intervalo para uma reflexão: por que estou escrevendo “protista” entre aspas? Esse grupo de seres eucariontes unicelulares eram considerados um Reino. Entretanto, hoje se sabe que os “protistas” não formam uma linhagem única, e podemos ver isso na Figura 2: se excluirmos plantas, animais e fungos, todos os outros nomes se referem a organismos que anteriormente eram incluídos nesse “Reino Protista”. Assim, continuamos usando esse termo por conveniência, mas não se refere a um agrupamento monofilético de organismos. Monofilético são os grupos que têm uma história evolutiva comum pois descendem do mesmo ancestral.
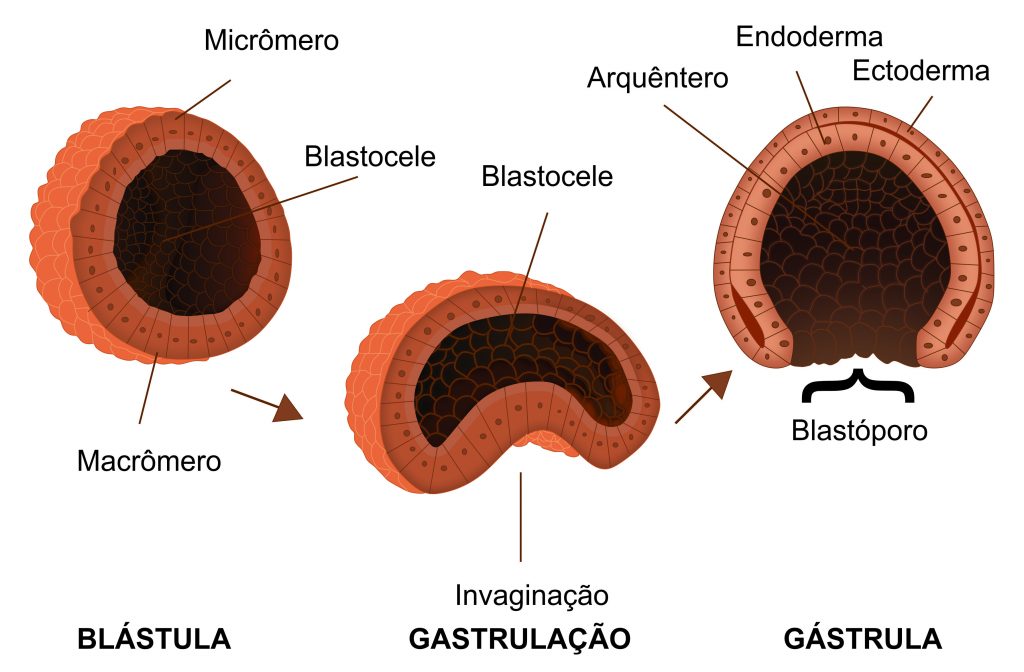
Os Metazoa são um grupo monofilético definido por muitas novidades evolutivas (= sinapomorfias; leia o texto “Como identificar relações de parentesco entre os seres vivos?”). Além da multicelularidade e da presença de junções epidérmicas aderentes, todos apresentam um estágio embrionário com blástula e todos, exceto os Porifera (popularmente chamados de “esponjas”), apresentam o estágio de gastrulação (Figura 3), que gera duas camadas embrionárias: epiderme externa e endoderme interna; a presença de diferentes camadas embrionárias leva à produção de estruturas distintas, aumentando a diversidade de formas dentro do grupo. Os animais possuem, ainda, processos únicos na produção de ovócitos (gametas femininos) e de espermatozóides (gametas masculinos), células com metade da carga genética do indivíduo adulto (= haplóides, símbolo “n”) que, quando se unem durante o processo de fecundação, resultam em um organismo com dois conjuntos de material genético, um vindo do ovócito da mãe e outro do espermatozóide do pai (= diplóide, símbolo “2n”). Apresentam, ainda, estruturas relacionadas à sustentação e movimentação corporal, como as proteínas contráteis actina-miosina e a proteína colágeno.
Ao final da era Proterozoica, há cerca de 750 milhões de anos, começou a quebra do supercontinente Rodínia. Nessa era ainda havia pouco oxigênio na atmosfera e nas regiões mais profundas do oceano. Há indícios da ocorrência de três períodos de glaciações, cada um durando cerca de 10 milhões de anos, separados por períodos de aquecimento forte e rápido, provavelmente produzido por fortes erupções vulcânicas que provocavam o acúmulo de CO2, resultando em um efeito estufa; nesse contexto surgiram os metazoários. Posteriormente, quando o nível de oxigênio na atmosfera aumentou e se estabilizou, teria permitido a rápida diversificação de organismos grandes e mais ativos.
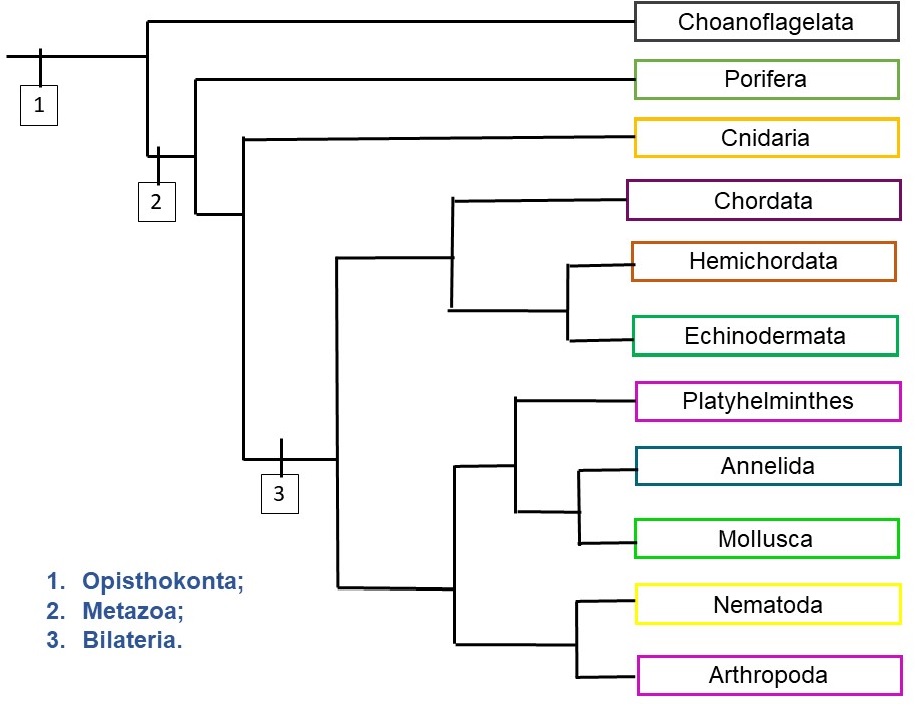
No cladograma resumido dos Metazoa (Figura 4) estão os nomes dos maiores filos animais, para que você tenha um panorama geral do Reino. Para cada ponto da história evolutiva desses organismos será citada, nesse texto, ao menos uma novidade evolutiva que a partir dali levou à grande diversificação das linhagens descendentes.
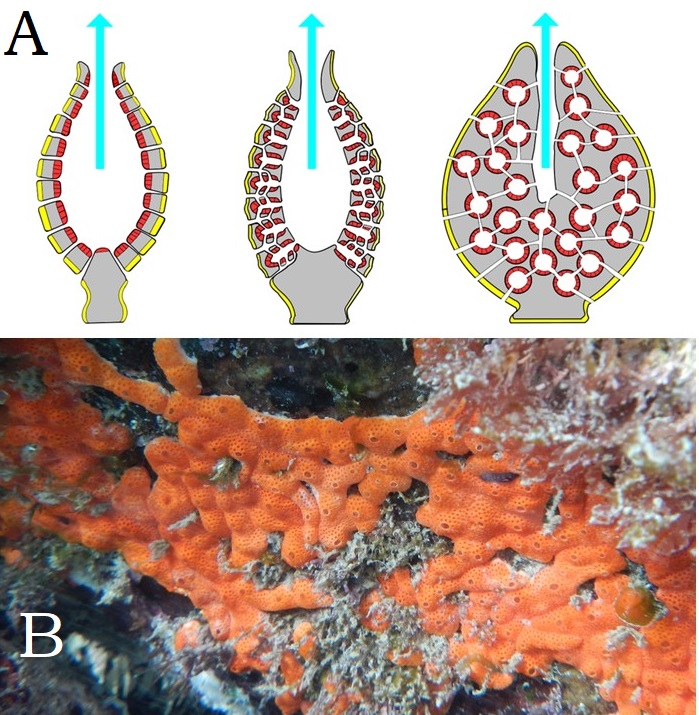
Embora seja difícil perceber o porquê dos exemplares incluídos no Filo Porifera serem considerados animais, eles compartilham conosco as características que definem Metazoa. Entretanto, não formam tecidos verdadeiros. Alimentam-se de partículas em suspensão capturadas pelos coanócitos, células flageladas muito semelhantes aos organismos coanoflagelados. A água entra em seus corpos por vários pequenos poros e sai por uma abertura maior, o ósculo; a parede corporal pode ser bastante elaborada, aumentando a área superficial em contato com a água, a quantidade de células corporais e potencializando a filtração e captura de partículas (Figura 5). A maior parte das espécies atuais é de água salgada, mas algumas espécies ocorrem em água doce. Muitos membros desse grupo apresentam esqueletos internos formados por espículas calcárias e silicosas, o que garante melhores condições de fossilização.
O filo Cnidaria se caracteriza por ser diblástico: como seu embrião sofre gastrulação, desenvolverá dois tecidos embrionários (Figura 6): a ectoderme externa e a endoderme interna. Muito diverso, inclui formas muito conhecidas, como medusas, anêmonas-do-mar e corais; esses últimos, que formam extensas colônias interligadas por seus esqueletos de carbonato de cálcio formam os mais importantes registros fósseis desse filo. Nesse grupo, radialmente simétricos, ocorre um sistema digestório incompleto, onde a ingestão de alimento e a eliminação de resíduos não digeridos (= fezes) ocorrem pela boca.

Os grupos seguintes são bilateralmente simétricos e triploblásticos: durante a neurulação, fase que sucede à gastrulação, a camada embrionária interna, endoderme, diferenciará um terceiro folheto germinativo, a mesoderme. Esse folheto embrionário é responsável pela origem de muitos tecidos e estruturas, como musculatura, tecidos conjuntivos, estruturas esqueléticas e sistemas circulatório, excretor e reprodutor. Em alguns grupos o peritônio, tecido mesodérmico, margeia uma cavidade corporal, o celoma; o sanduíche formado pela bolsa de peritônio com celoma interno, preenchido por líquido celomático, fica em volta dos órgãos corporais. Notamos, portanto, como o surgimento da mesoderme é fundamental para a origem da crescente complexidade corporal que observamos nos animais.
O filo Platyhelminthes inclui vermes achatados dorsoventralmente e acelomados, pois neles a mesoderme não forma cavidade celomática. Há espécies de vida livre em água doce ou salgada, mas a maioria delas é de parasitas; tubo digestório incompleto e até mesmo ausente em algumas formas. Seus corpos moles não formam registros fósseis importantes.
Os moluscos são caracterizados por possuírem corpo com cabeça, pé e massa visceral, essa última coberta por um tecido chamado “manto” que geralmente secreta o esqueleto em forma de concha, composto por carbonato de cálcio (Figura 7). Alguns grupos não têm concha, porém, ou têm concha reduzida e interna. Entretanto, graças à essa estrutura, formam um rico e informativo registro fóssil. Os Anelídeos, grupo proximamente relacionado, têm corpo fortemente segmentado, muitas formas com o corpo repetindo várias vezes segmentos muito parecidos entre si, geralmente com cerdas, e sistema nervoso ganglionar bem desenvolvido. Animais com essas características são conhecidos aproximadamente a partir do Cambriano.

Nas rochas do período Ediacarano (final da era Proterozoica) nos deparamos com uma grande surpresa: há registros de invertebrados marinhos que incluem formas muito parecidas à de filos que ainda ocorrem hoje em dia, como os Porifera (esponjas), Cnidaria (corais e águas-vivas), Mollusca (caramujos e polvos). Alguns dos mais de 100 gêneros de fósseis ediacaranos descritos tanto de águas rasas quanto profundas, porém, são diferentes de tudo o que conhecemos. Aparentemente a maioria deles possuía corpos moles, sem partes duras de grandes tamanhos. Já se encontra registros de duas formas corporais: os que apresentavam simetria radial, como os Cnidaria e equinodermas atuais, e os com simetria corporal bilateral. Muitos desses organismos entraram em extinção na transição do para o Cambriano, no último evento de glaciação do Proterozoico.

O éon Fanerozoico, que inclui as eras Paleozoica, Mesozoica e Cenozoica – essa última sendo a nossa era atual – é palco de um período de ocorrência de muitas formas de vida. Começou, na era Paleozoica (541 a 252 Ma), com a “explosão do Cambriano” (Figura 8), período entre 541 e 485 Ma no qual os animais continuaram a enfrentar oscilações climáticas extremas e mudanças químicas da atmosfera e dos oceanos. O aumento no teor de cálcio dos mares influenciou o surgimento de muitos organismos com esqueletos bem-mineralizados inclusive representantes da maior parte dos filos animais que conhecemos atualmente. Talvez a “popularização” desses esqueletos seja um indício de que nesse período aumentaram as atividades predatórias nos mares. Enquanto a fauna ediacarana inclui maior número de espécies que se alimentavam de detritos e material em suspensão, no começo do Cambriano há registros de carnívoros e herbívoros, como artrópodes predadores gigantes, e no final do Cambriano já ocorrem registros de peixes agnatos, talvez hematófagos (comedores de sangue) ou saprófagos (comedores de animais mortos) como os que existem atualmente. Assim, ampliaram-se muito os nichos ecológicos nos ambientes marinhos, e muitas dessas linhagens cambrianas viveram por muitos milhões de anos.
Trabalhos publicados recentemente sugerem que alguns desses filos tenham, na verdade, se originado no período Ediacarano. Algumas das discussões avaliam a taxa de oxigênio livre nesses períodos, que certamente está ligada à evolução da vida animal. Provavelmente foram necessários alguns milhões de anos para acúmulo do oxigênio livre produzido pela fotossíntese das cianobactérias. É possível, também, que os oceanos proterozoicos tenham sido mais oxigenados nas regiões mais superficiais e mais anóxicos nas regiões mais profundas. O oxigênio permitiu a evolução de formas maiores e de formas mais ágeis, que podiam sair em busca de suas presas, gastando muita energia nesse processo.
Nematoda e Artropoda formam, com outros filos menores, o clado Ecdysozoa, pois trocam a cutícula quitinosa ao menos uma vez durante seu ciclo de vida. Nematoda geralmente têm uma troca entre cada um dos 4 estágios de crescimento. Apresentam um par de órgãos sensoriais cefálicos, os anfídios, que consistem em um poro externo ligado por um ducto a uma bolsa anfidial ligada ao anel nervoso cerebral; provavelmente são quimiorreceptores. Os Artrópodos são, de longe, o maior grupo de animais com espécies descritas, e com seu exoesqueleto quitinoso estão presentes em registros fósseis desde o pré-Cambriano. Ainda assim, estima-se que o que conhecemos é cerca de 10 a 20 % de todas as formas que existem.

O modo de formação da boca, ânus e celoma é um dos critérios que agrupa os animais dos filos Hemichordata, Echinodermata e Chordata como deuterostomados. Há uma proposta de que os dois primeiros filos sejam mais próximos entre si por compartilharem o mesmo destino do blastóporo, que geralmente forma ânus (embora alguns grupos tenham sofrido regressão e apresentem tubo digestório incompleto) e por exibirem uma larva com estrutura bem parecida. Ambos os grupos só têm representantes marinhos. Os Hemichordata são vermes de corpo mole dividido em probóscide ou escudo, colarinho e tronco. Apresentam uma estomocorda que já foi considerada homóloga da notocorda, embora a opinião atual é de que não seja; daí o seu nome. Já os Equinodermas, com simetria bilateral nas larvas e simetria radial desenvolvida secundariamente no adulto, têm um complexo sistema de canais que forma um sistema ambulacral que, além de auxiliar na locomoção, serve também para respiração, excreção e captura de alimento, a depender do organismo. Equinodermos também apresentam um rico registro fóssil, já que a maioria das formas apresenta um endoesqueleto de carbonato de cálcio.

Os Chordata são um grupo mais conhecido por incluírem os Vertebrata. Atualmente passa por muitas modificações de nomenclatura resultantes da metodologia cladista. O grupo “Pisces”, por exemplo, não é monofilético e, na verdade, engloba quatro grupos principais: Myxini (feiticeiras), Petromyzontiformes (lampréias), Chondrchthyes (‘peixes’ secundariamente cartilaginosos) e Osteichthyes, os peixes ósseos. Esse último inclui dois grupos principais: os Actinopterygii, que inclui cerca de 30 mil espécies de ‘peixes’ nos quais a nadadeira só apresenta raios, sem musculatura, e Sarcopterygii, com 8 espécies de ‘peixes’ ainda existentes e os Tetrapoda. Os Tetrapoda incluem os Anfíbios e os Reptilomorpha. Nesse último grupo estão incluídos os répteis, aves e mamíferos. Prestou atenção no que está escrito aqui? Nós, mamíferos, somos um tipo de Osteichthyes, portanto…

Os primeiros peixes teleósteos (grupo de Actinopterygii onde são incluídas a maioria das espécies que ocorrem no planeta atualmente) e os famosos euriptéridos (conhecidos como “escorpiões-marinhos”, embora não sejam escorpiões), proliferaram no período Ordoviciano. Os primeiros artrópodos terrestres surgiram no Siluriano e no Devoniano o número de formas de vida terrestres aumentou muito, graças à diminuição do CO2 atmosférico (veja no texto sobre “Os vegetais”). Cerca de 80% das espécies conhecidas pelos cientistas atualmente são artrópodes, e os insetos são cerca de 80% desse total.
No Permiano há cerca de 270 Ma, houve a maior extinção em massa do Planeta, na qual cerca de 90% das espécies marinhas e 70% das terrestres foram eliminados, incluindo formas comuns até então, como alguns tipos de corais e os trilobitas.
No Triássico (já na era Mesozoica, há cerca de 200 Ma) houve a formação da Pangeia. O clima do planeta era quente, e haviam grandes desertos. Após a extinção de Permiano a diversidade de animais ampliou-se rapidamente, e surgiram os dinossauros e os primeiros mamíferos terrestres. Ao menos na parte mais rasa dos mares sabemos que aumentou a diversidade de corais e peixes. Houve, porém, um novo evento de extinção em massa, que resultou no desaparecimento de aproximadamente metade das espécies da época.

O período Jurássico continuou com clima quente. Nesse período a Pangeia dividiu-se em Laurásia ao norte e Gonduana ao sul, gerando um canal marinho, o Mar de Tétis, que parece ter sido ambiente para muitas espécies da fauna marinha tropical.
Os oceanos, inclusive o Atlântico, sofreram muitas transgressões: as águas, mais altas, avançavam sobre as terras, mais baixas, formando muitas áreas aquáticas rasas. Aos poucos o clima foi esfriando. Ao final do período Cretáceo, nova extinção em massa, com perda de cerca de 50% das espécies, inclusive muitas formas de dinossauros todos os amonóides. O Cenozoico, que começou há 66 Ma, continuou com clima mais frio por cerca de 10 milhões de anos. As correntes marinhas que se formaram quando a Antártida se separou da América do Sul resultou no seu clima gelado atual. Há 56 Ma os gases do efeito estufa aqueceram a atmosfera e os oceanos do planeta e várias linhagens de animais voltaram a aumentar, como os recifes de corais atuais. Agora, discute-se sobre o rápido declínio e extinção de muitas formas de vida causada pelo intenso crescimento das populações humanas e suas atividades econômicas em diversos pontos da Terra.