

Por Rosana Souza-Lima
Você já deve se perguntou por que alguns biólogos se preocupam tanto em catalogar as diversas espécies de seres vivos do planeta? Por que será que é importante saber em que grupo uma determinada espécie se encaixa? A área de trabalho que lida com essas questões é chamada de “Sistemática”, justamente pela preocupação em sistematizar (=organizar) nossos conhecimentos sobre os seres vivos. A sistemática, porém, vai muito além de apenas conhecer ou listar as espécies conhecidas. Vamos conversar sobre isso!
Imagine que queiramos identificar, por exemplo, os nichos alimentares que podem ser encontrados em determinada área, ou quais as relações ecológicas entre dois ou mais seres vivos de um dado lugar ou até mesmo comparar as proteínas que ocorrem em um determinado grupo de espécies. Bom: a primeira coisa que precisamos descobrir é de que espécies estamos falando, não é?
Nossas tentativas de reconhecer espécies e os modos de agrupá-las são muito, muito antigas: começaram cerca de 350 anos antes da Era Cristã, com os trabalhos do filósofo Aristóteles. A partir daí alternativas se sucederam, sendo mais ou menos aceitas pela comunidade científica. Em 1950 o entomólogo alemão Willi Hennig apresentou uma proposta metodológica que ganhou espaço ao longo dos anos: a metodologia cladista. Como funciona esse método?
Para que possamos avaliar os caracteres do ponto de vista evolutivo devemos, de início, formular uma primeira hipótese: serão esses caracteres homólogos ou homoplásicos? Homólogos são caracteres que possuem a mesma origem evolutiva, o que indica, portanto, parentesco entre os grupos comparados. As homoplasias são características muito parecidas entre si, mas que não têm uma mesma origem evolutiva. Ou seja: os caracteres ficaram parecidos por processos que os modelaram em resposta a alguma pressão ambiental. Vamos examinar um exemplo: à primeira vista, as nadadeiras de peixes e baleias são muito parecidas entre si. Entretanto, uma simples radiografia mostra que são formadas por estruturas muito diferentes: a semelhança é apenas externa, causada pelo fato de ser mais fácil se locomover no ambiente aquático possuindo uma estrutura que empurre a água. Além da aparente semelhança entre as estruturas comparadas, a posição anatômica em que essas estruturas se situam pode dar uma pista sobre sua provável homologia. E se for possível estudar o desenvolvimento embrionário dos organismos em questão, melhor ainda, pois os organismos mais relacionados entre si são muito parecidos durante o início do seu desenvolvimento. Quanto mais evidências analisamos, mais confirmamos a nossa hipótese sobre “aquele” ser um caráter que nos ajudará a identificar o parentesco entre os grupos estudados.
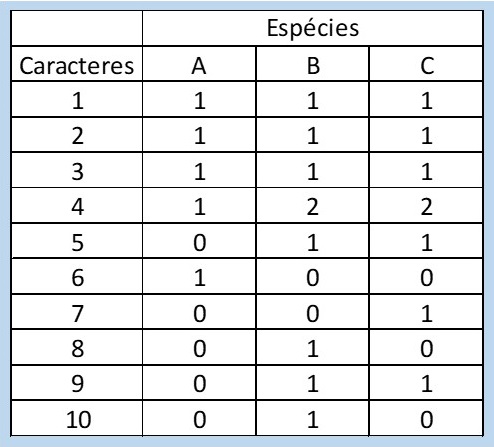
Uma vez definidas as homologias, chegamos ao ponto principal: selecionar quais delas são importantes na indicação do relacionamento entre as linhagens. Essa avaliação é o “pulo do gato” nessa metodologia! Como fazemos isso? Uma das possibilidades é comparar o grupo que estamos estudando (que agora chamaremos de grupo interno) com algum outro grupo que seja parecido com ele (grupo externo). O fato de serem parecidos facilitará na hora de compararmos as estruturas de ambos, porque nem sempre é fácil entendermos se se trata da mesma coisa. A regra é simples: se a característica examinada se apresentar da mesma forma em ambos os grupos, ela é considerada uma plesiomorfia: significa que é um caráter que já estava presente no ancestral de todos, e não representa uma novidade evolutiva para os grupos que estão sendo avaliados. Já se um dado caráter for diferente em um, alguns ou em todos os membros do grupo interno quando comparados aos membros do grupo externo, o caráter é considerado uma novidade evolutiva ou, em “biologuês”, uma apomorfia. Essa avaliação é a “polarização de caracteres”: a partir dessas deduções construiremos um quadro (=matriz) onde as plesiomorfias serão codificadas com zero, e as apomorfias codificadas com um; zero e um são os estados do caráter. Apenas as apomorfias, portanto, indicam parentesco entre os membros do grupo interno, que é o nosso grupo de interesse; o grupo externo só é utilizado para nos permitir formular hipóteses sobre a natureza dos caracteres que estamos observando. Por exemplo, vamos construir uma matriz a partir da comparação dos peixinhos exibidos na Figura 1: um desses taxa representa o grupo externo, e os 3 taxa seguintes representam espécies distintas (“Táxon” é uma palavra de origem latina que significa qualquer unidade taxonômica: pode ser usada para designar espécie, família, ordem, tribo… qualquer categoria! Seu plural é “taxa”).
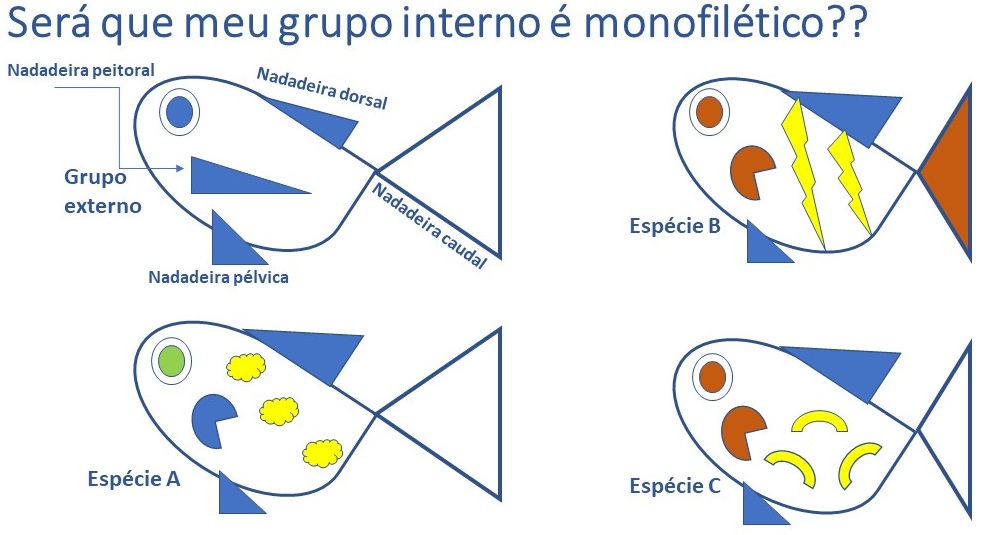
O primeiro passo será levantarmos características a serem examinadas por comparação com o grupo externo. Ex.:
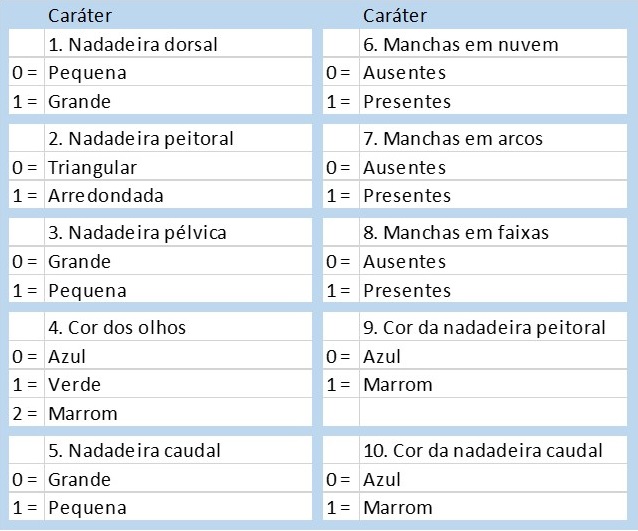
A partir das polarizações que estamos propondo, construímos nossa matriz. Nessa matriz nem colocamos uma coluna para o grupo externo, porque como regra da metodologia já sabemos que todos os caracteres que ele possui são polarizados como plesiomorfias. Perceba que não adianta, portanto, computarmos as características que permanecem iguais às que ocorrem no grupo externo: essas características, como formato do corpo e formato do olho, são plesiomorfias que ocorrem em todos os 4 grupos e, portanto, não indicam parentesco. Um caráter plesiomórfico que une vários grupos é chamado de simplesiomorfia (sin = ‘junto’), e não tem valor informativo porque interpreta-se que já ocorria no ancestral dos grupos externo e interno. As novidades evolutivas, ou apomorfias, quando unem taxa são chamadas de sinapomorfias. Quantas sinapomorfias estão listadas na tabela abaixo?
De posse da nossa matriz, vamos construir uma representação gráfica do processo evolutivo dessa linhagem que é chamado de cladograma. O cladograma começa mostrando uma linha do tempo, onde aos poucos vamos incluindo os fatos da história que estamos contando. Podemos começar pelos fatos mais abrangentes, que reúnem um maior grupo de taxa, para os menos abrangentes, que atinge apenas um táxon. Por exemplo: os caracteres 1, 2 e 3 ocorrem em todas as três espécies. Então, a história que contam começa assim:
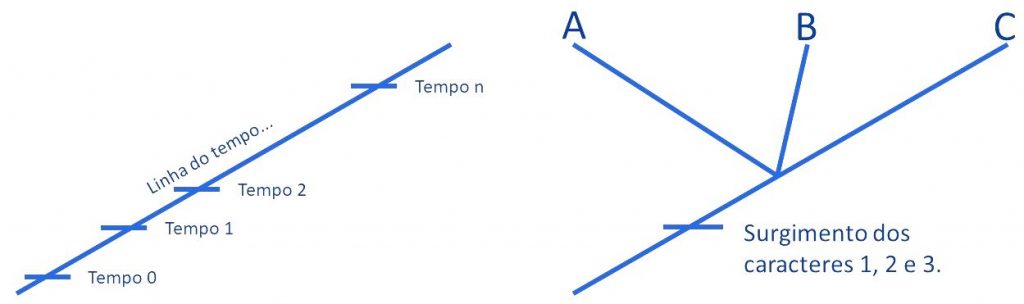
O que significa a história contada na Figura 2? Que em determinado momento, ou em 3 momentos distintos, as alterações genéticas que se acumulavam numa dada linhagem resultaram no surgimento de 3 caracteres diferentes dos que já existiam. Nesse momento não conseguimos distinguir os indivíduos A, B e C entre si, porque todos os 3 tinham as 3 características novas. Essas características novas, portanto, são sinapomorfias que unem A, B e C. Sendo unidos por sinapomorfias, podemos dizer que o grupo ABC é monofilético: possui apenas uma (=mono) origem (=filético); o ancestral das linhagens ABC é o mesmo. Mas o que aconteceu depois? Como foi a diferenciação seguinte?
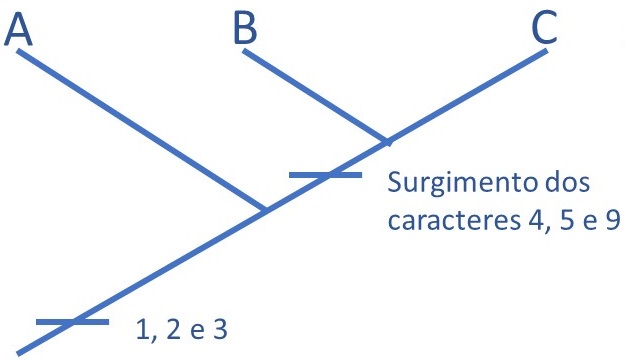
Os caracteres 4, 5 e 9 nos fornecem a próxima pista: B e C formam um segundo grupo monofilético (Figura 3). O compartilhamento desses três caracteres mostra que B e C são mais próximos entre si do que do A. Podemos perceber, ainda, a plasticidade dos caracteres: enquanto os caracteres 1, 2 e 3 eram uma sinapomorfia que reúne A, B e C, eles podem ser encarados como uma simplesiomorfia para B e C. Ou seja: eles não servem mais para agrupar B e C, porque o surgimento de 1, 2 e 3 ficou para trás, para um outro momento dessa história. Foi necessário o surgimento de novas apomorfias, 4, 5 e 9, para que percebêssemos que B e C formam um grupo monofilético.
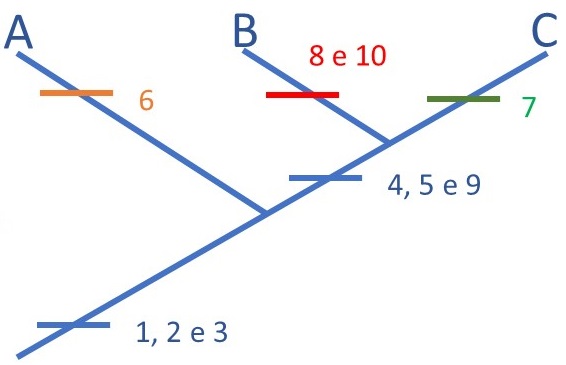
Para que servem os caracteres escritos em cores diferentes do azul na Figura 4? Esses caracteres que são exclusivos de um táxon, e que não unem dois grupos, são chamados de autapomorfias. Embora não sejam úteis, nesse momento das nossas análises, para esclarecer as relações de parentesco entre ABC, servem para diagnosticar cada um desses taxa. No campo, por exemplo, se encontrarmos um peixinho desse tipo com manchas amarelas em formato de nuvens, já saberemos que se trata de um exemplar da espécie A. A presença dos caracteres autapomórficos nos mostra, também, que a todo momento estão surgindo modificações nas características de uma linhagem. Essas modificações são causadas por influência ambiental, ao longo do tempo, na carga genética dos organismos, provocando alterações que aos poucos se acumulam. Essa diferenciação genética ao longo do tempo se chama anagênese, e cria as condições gênicas para que aos poucos as espécies se diferenciem. Já falamos sobre anagênese e cladogênese na publicação sobre “Fósseis e a evolução dos seres vivos”. Dê mais uma olhada lá para relembrar!
Nem sempre os caracteres apomórficos apresentam-se em apenas um estado: você pode registrar todas as variações que vê em uma determinada estrutura. Já está implícito na metodologia que a ordem em que alguém os numera não necessariamente é a ordem em que apareceram na natureza. Pode inclusive ficar comprovado que apareceram em completa desordem. Veja um exemplo na Figura 5! No nosso grupo hipotético de peixes também temos um caráter multiestado, o caráter número 4 (Tabela 2). Veja que nesse caso não precisamos de nenhuma hipótese de ordenação dos caracteres para que ele nos fornecesse uma informação de parentesco entre os grupos B e C (Figura 4).
De acordo com a composição dos grupos internos e externos podemos polarizar o caráter de um modo diferente. Assim, como praticamente tudo na Ciência, as classificações estão sujeitas a novos ajustes e refinamentos à medida que novos taxa e novos caracteres forem acrescentados. E a partir de estudos das relações de parentesco entre organismos também temos uma ideia que corroboram ou não propostas geológicas e geográficas de como a Terra foi se modificando ao longo desses milhões de anos em que a vida se transforma. É uma linha de trabalho fascinante!
ótimo texto, muito bem escrito e extremamente esclarecedor sobre como funciona a sistemática cladista.